Дифтерийный токсин
Cтраница 1
Дифтерийный токсин представляет собой белок с мол. Его минимальная летальная доза для морской свинки составляет всего лишь 0 16 мг / кг. Исследования, проведенные на культуре клеток, показали, что токсин блокирует включение аминокислот в белки в результате инактивации-фактора элонгации EF-2, необходимого для транслокацин в рибосомах млекопитающих. [1]
Дифтерийный токсин очищают при помощи фракционированного осаждения сернокислым аммонием. Дифтерийный токсин представляет собой глобулиноподобный белок, разрушающийся при нагревании и под действием протеоли-тических ферментов. [2]
Иммунитет клеток к дифтерийному токсину также связан с мембранными барьерами. Этот микробный яд последовательно поражает два типа молекулярных мишеней в животных клетках. Один находится в составе клеточной мембраны, второй располагается внутри клетки. Структура мембранной мишени первого типа и обусловливает невосприимчивость или уязвимость животных клеток к токсину. Какое-то пока еще не известное преобразование ее задерживает проникновение дифтерийного яда внутрь клетки. [3]
Недавно установлено химическое строение дифтерийного токсина. Он представляет собой сложный эфир миколо-вой кислоты и дисахарида трегалоэы. Помимо описанных возбудителей туберкулеза, проказы, дифтерии, есть и другие виды микобактерий, патогенные для человека и животных. Фитопатогенные формы встречаются редко. [4]
В состав молекулярной мишени дифтерийного токсина входят уже совсем другие вещества, а именно фосфо-липиды клеточной оболочки, причем присоединение ток-синной молекулы к фосфолипидам осуществляется ее гидрофобным С-кольцом. После присоединения к поверхности клеточной мембраны молекула дифтерийного токсина каким-то образом разделяется на два фрагмента с массой 24 000 и 34 000 дальтон соответственно. Больший фрагмент остается на клеточной оболочке. Он служит только для прикрепления к последней, и его функция на этом заканчивается. Меньший фрагмент проникает сквозь оболочку внутрь клетки, где находится конечная мишень его поражающей активности. [5]
Наиболее изученный из них - дифтерийный токсин. [6]
Весьма интересен молекулярный механизм действия дифтерийного токсина. Он оказался наделенным способностью катализировать реакцию АДФ-рибозилирования фактора элонгации эукариот ( eEF - 2), выключая тем самым его из участия в синтезе белка. Резистентность многих животных к дифтерийному токсину, вероятнее всего, обусловлена трудностью или полным отсутствием проникновения ( транспорта) токсина через мембрану клеток. [7]
Клетки животных становятся иммунными к дифтерийному токсину и в том случае, если их обработать ганглиозидами определенных видов. Но после введения в организм животного других гаиглиозидов возникает иммунитет к столбнячному токсину. При этом предотвращается взаимодействие токсина с теми структурами нервных клеток, которые он обычно поражает. [8]
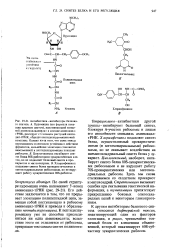
К другим ингибиторам белкового синтеза относятся дифтерийный токсин, инактивирующий один из факторов элонгации, и рицин, чрезвычайно токсичный белок из клещевины обыкновенной, который инактивирует 608-суб-частицу эукариотических рибосом. [10]
Необходимо отметить, что исходные молекулы дифтерийного токсина не обладают таким ингибиторным действием на белковый синтез; они скорее являются зимогеном, превращаемым в каталитически активный белок ( А-фрагмент) лишь после расщепления. С другой стороны, сам фрагмент А не обладает цитотоксическим действием, так как не может проникнуть в интактную клетку. [11]
Клетки человека очень чувствительны к поражающему действию дифтерийного токсина. И тем не менее некоторые кз них в полной мере сохраняют жизнеспособность. Такую же иммунность проявляют и их потомки. Какие-то наследственные силы помогают им спастись. [12]
Несмотря на знание многих деталей процесса превращения дифтерийного токсина в энзиматически активный фрагмент А и энзимати-ческого действия фрагмента А на EF-2, механизм происходящей в результате инактивации белкового синтеза остается загадкой. Дело в том, что АДФ-рибозилирование не приводит, как ожидалось, к инактивации известных парциальных функций фактора. АДФ-рибо-зилированный EF-2 сохраняет способность взаимодействовать с ГТФ как эффектором, и комплекс ADPR-EF - 2 GTP связывается с рибосомой так же, как и исходный EF-2 GTP. Связывание происходит по тому же рибосомному участку. ГТФазная активность тоже обнаруживается как результат взаимодействия модифицированного фактора ( плюс ГТФ) с рибосомой. Правда, в ряде работ отмечалось, что в случае АДФ-рибозилированного EF-2 ГТФазная активность может быть в той или иной степени снижена и что для промотирования транслокации требуются не каталитические, а стехио-метрические количества фактора по отношению к рибосомам. Были высказаны предположения, что ингибирование белкового синтеза в результате АДФ-рибозилирования EF-2 может достигаться либо за счет понижения сродства модифицированного EF-2 к факторсвязываю-щему участку рибосомы, либо за счет уменьшения скорости его реутилизации ( recycling), например, вследствие малой скорости диссоциации комплекса рибосома ADPR-EF - 2 GDP. Не понятным, однако, остается тот факт, что АДФ-рибозилирование EF-2 приводит к полной остановке белкового синтеза, в то время как парциальные функции EF-2 если и нарушаются, то лишь до некоторой степени. [13]
Травма роговицы плотными пленками, ядовитое действие дифтерийного токсина, нередкое наслоение гноеродной инфекции приводит иногда к развитию кератита и к гнойному расплавлению роговицы с последующим образованием бельма и понижением зрения, вплоть до слепоты. [14]
Она появляется в ответ на введение растворимых белковых антигенов ( дифтерийный токсин, пыльцовые, эпидермальные аллергены) или конъюгатов через 6 - 8 ч после внутрикожной реакции и стихает через 1 - 2 сут. Внешне проявляется эритемой и припухлостью, на срезах кожи видна лимфоидная инфильтрация вокруг мелких сосудов и капилляров. Реакция Джонса Мота исчезает по мере накопления в крови циркулирующих антител. [15]
Страницы: 1 2 3 4