Инициация - репликация
Cтраница 1
Инициация репликации строго регулируется. Полирепликонная организация требует, чтобы в каждом цикле клеточного деления каждый ориджин сработал только один раз, в противном случае на хромосоме образуются разветвленные структуры. [1]
Инициация репликации ДНК в клетках эукариот регулируется на уровне кластера репликонов, причем число репликонов в кластере изменяется от нескольких единиц до нескольких десятков. [2]
Инициация репликации ДНК в клетках млекопитающих возобновляется, начиная примерно с 2 ч после облучения их в дозе несколько Грей [ Walters R. К этому времени ДНК должна находиться в суперспиралыюй конфигурации [ Mattern M. R., Painter R. К 2 ч после облучения во фракции более радиорезистентных тимоцитов эксцизионная репарация еще не завершается ( см. рис. 13), что не позволяет возобновиться инициации репликации до момента полного воссоединения разрывов и восстановления суперспиральной: конфигурации ДНК. В то же время во фракции радиочувствительных клеток к 1 - 2ч после облучения воссоединение разрывов уже произошло, но инцизия еще не наступила. Возобновление репликации к этому времени в дальнейшем может привести к фрагментации ДНК за счет образования разрывов инцизии в однонитевых участках, примыкающих к репликативной вилке и возникающих в пределах репликона на пути роста цепи. [3]
Инициация репликации ДНК требует, чтобы к сайту начала репликации присоединились несколько молекул белка DnaA. Активная форма этого белка связана с АТФ, поэтому регуляция инициации может осуществляться взаимопереходом DnaA-АТФ и DnaA-АДФ. Следующий цикл репликации начинается не сразу, так как родительская нить ДНК метилируется сразу после репликации и становится неактивной. [4]
Другие способы инициации репликации целесообразно рассмотреть в связи с вопросом о регуляции репликации. Молекулярные-основы подобной регуляции лучше всего изучены в случае некоторых бактериальных плазмид. [5]
Другие способы инициации репликации целесообразно рассмотреть в связи с вопросом о регуляции репликации. Молекулярные основы подобной регуляции лучше всего изучены в случае некоторых бактериальных плазмид. [6]
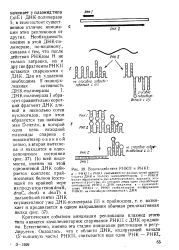 |
Взаимодействие РНКП и PHKI. [7] |
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКП с ДНКориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [8]
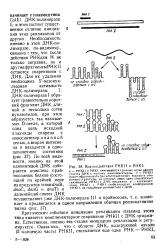
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКИ с ДНК ориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [10]
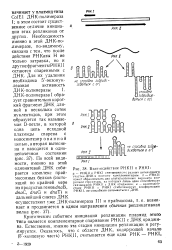 |
Взаимодействие РНКП и PHKI. [11] |
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКП с ДНКориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [12]

Одни плазмиды несут специфичный сайт инициации репликации и могут реплицироваться только в клетках одного вида. У других плазмид этот сайт менее специфичен, и они реплицируются в самых разных бактериальных клетках. Соответственно различают плазмиды с узким и с широким спектром хозяев. [14]
 |
Фрагмент реплицирующейся ДНК хромосомы из разрушенных ядер Dro. [15] |
Страницы: 1 2 3 4