Инициация - трансляция
Cтраница 1
Инициация трансляции представляет собой импульс, сообщаемый молекулярной машине, ориентированной на биосинтез белка. [1]
Инициация трансляции в про - и эукариотических клетках имеет много общих черт. [2]
В некоторых редких случаях инициация трансляции может происходить на AUG триплете, отстоящем довольно далеко от 5 -конца. Например, в мРНК эпидермального фактора роста мышей инициирующий кодон отнесен от 5 -конца на 350 нуклеотидных остатков, но и в этом случае он - первый AUG триплет от конца. Имеются также другие редкие примеры, где в не транслируемых 5 -концевых последовательностях встречаются AUG триплеты, проскакиваемые рибосомами без инициации; очевидно, они не могут служить инициаторами из-за отсутствия подходящего структурного окружения. Иногда на сравнительно небольшом расстоянии вслед за AUG триплетом следует терминирующий кодон, который, по-видимому, предотвращает его использование в качестве инициатора синтеза длинного полипептида. Однако в случаях некоторых вирусиндуцированных мРНК может наблюдаться такое явление, когда первый AUG триплет узнается как инициирующий лишь частью рибосом ( слабый инициатор), в то время как остальная часть сканирующих рибосом его проскакивает без инициации и инициирует на следующем AUG; тогда мРНК оказывается функционально двуцист-ронной, так как на ней начинается и происходит синтез двух разных полипептидов, кодируемых перекрываемой нуклеотидной последовательностью. [3]
Результаты анализа функциональных сайтов инициации трансляции представлены в таблице зь. [4]
РНК фиксируется в нужном для инициации трансляции положении. [5]
Итак, во всех случаях инициации трансляции предшествует диссоциация нетранслирующих 70S рибосом на 30S и 50S субчастицы. Как уже говорилось, в физиологических условиях нетранслирующие рибосомы находятся в обратимом равновесии со своими субчастицами. [6]
Необходимо отметить, однако, что инициация трансляции свободными рибосомами в какой-то мере возможна, по-видимому, также и на матрицах без последовательности Шайна - Дальгарно. X, транскрибируемая с промото - Ра - ркм ( в лизогенной клетке), вообще не имеет прединициирующей последовательности, но инициация на ней происходит, хотя и не очень эффективно. Бактериальные рибосомы могут осуществлять также инициацию на некоторых чужеродных ( эукариотиче-ских) РНК, лишенных последовательности Шайна - Дальгарно, например, на РНК 4 вируса мозаики костра; инициация требует всех трех бактериальных факторов инициации и происходит на нормальном инициирующем кодоне. Отсюда последовательность Шайна - Дальгарно, безусловно, вносит важный вклад в эффективную ассоциацию рибосомных частиц с матрицей, но не является абсолютным требованием. [7]
Вероятность перехода той же рибосомы к инициации трансляции следующего цистрона определяется, по-видимому, длиной и структурой меж-цистронового участка: чем он короче и менее структурирован, тем больше шансов на инициацию без диссоциации. Однако и в данном случае похоже на то, что на матрице удерживаются лишь 30S субчастицы, a 50S субчастицы, как результат терминации трансляции, находятся с ними в обратимом равновесии и, таким образом, обмениваются. [8]
Все это обсуловливает целый ряд особенностей инициации трансляции и ее регуляции у эукариот. [9]
Различия в механизмах трансляции в основном касаются процессов инициации трансляции. [10]
В целом картина с белковыми факторами, участвующими в инициации трансляции у эукариот, получается сложной. [11]
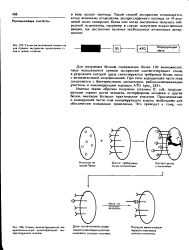 |
Схема расположения элементов для прямой экспрессии чужеродных re нов в новом хозяине.| Схема, демонстрирующая им-мунохимичеькую идентификацию экс-прессирующих клонов. [12] |
Присоединение к кодирующей части гена инициирующего кодона необходимо для обеспечения инициации трансляции. [13]
Оказалось, что в первичной ассоциации с мРНК и следующей затем инициации трансляции участвует, скорее, 30S субчастица, а не 70S рибосома. Именно 30S субчастица уводится из этой равновесной смеси на мРНК и затем вовлекается в первые стадии инициации. Факторы инициации - прежде всего IF-3 ( а также, возможно, IF-1) - способствуют сдвигу равновесия в сторону диссоциации свободных нетранслирующих рибосом на субчастицы. [14]
В связи со сказанным, существует специальный, довольно сложный механизм инициации трансляции, обеспечивающий как точное узнавание начала кодирующей последовательности и ее дальнейшую фазировку, так и возможность положительных и отрицательных регуляторных влияний. [15]
Страницы: 1 2 3 4