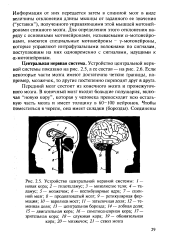
А-мотонейрон
Cтраница 1
Аксоны а-мотонейронов характеризуются высокрД, скоростью проведения - 70 - 120 м / с, что согласуется с их значительным диаметром. [1]
Как и а-мотонейроны, у-мотонейроны расположены в девятой пластине, где они рассеяны среди а-мотонейронов. Гамма-мотонейроны представляют собой значительно более мелкие клетки. Диаметр их тела не превышает 30 - 40 мкм. В отличие от а-мотонейронов у-мотонейроны не имеют непосредственного синаптического контакта с первичными афферентными волокнами. [2]
Деполяризация, возникающая в а-мотонейронах при активации возбуждающих синаптических входов, вызывает ритмические разряды потенциалов действия. Частота импульсации пропорциональна степени деполяризации, однако обычно она не превышает 10 - 20 имп / с, что обусловлено интенсивной следовой гиперполяризацией, развивающейся после каждого потенциала действия. [3]
Как и а-мотонейроны, у-мотонейроны расположены в девятой пластине, где они рассеяны среди а-мотонейронов. Гамма-мотонейроны представляют собой значительно более мелкие клетки. Диаметр их тела не превышает 30 - 40 мкм. В отличие от а-мотонейронов у-мотонейроны не имеют непосредственного синаптического контакта с первичными афферентными волокнами. [4]
Альфа-мотонейроны представляют собой крупные клетки с длинными дендритами. На соме и дендритах а-мотонейронов размещается до 10 000 - 20 000 синапсов, большая часть которых образована окончаниями интраспинальных вставочных нейронов. Часть синапсов на а-мотонейр-онах представляет собой контакты с окончаниями первичных афферентных волокон, идущих от мышечных рецепторов растяжения, и с волокнами длинных нисходящих трактов: кортико-спинального, руброспинального, вестибулоспинального и ретикулоспинального. [5]

Несмотря на то что разЙ чйые вставочные нейроны значительно отличаются друг от друга размерами, характером дендритных разветвлений, аксональными проекциями и функциональными особенностями, им присущи и некоторые общие свойства. Как правило, вставочные нейроны имеют тело меньшего диаметра, чем а-мотонейроны. Следовая гиперполяризация в большинстве разновидностей вставочных нейронов выражена незначительно. Поэтому они способны к генерации высокочастотной импульсной активности. Как видно на рис. 79, частота импульсации вставочного нейрона значительно превышает частоту импульсации мотонейронов. Даже в ответ на одиночный стимул они обычно генерируют множественные разряды. Частота импульсов, регистрируемых в теле некоторых вставочных нейронов, может превышать 1000 в секунду. [7]
Информация от них передается затем в спинной мозг в виде величины отклонения длины мышцы от заданного ее значения ( уставки), полученного управляющими этой мышцей мотонейронами спинного мозга. Для определения этого отклонения наряду с основными мотонейронами, называемыми а-мотонейро-нами, имеются специальные мотонейроны - у-мотонейроны, которые управляют интрафузальными волокнами по сигналам, поступающим на них одновременно с сигналами, идущими к а-мотонейронам. [9]
Как и а-мотонейроны, у-мотонейроны расположены в девятой пластине, где они рассеяны среди а-мотонейронов. Гамма-мотонейроны представляют собой значительно более мелкие клетки. Диаметр их тела не превышает 30 - 40 мкм. В отличие от а-мотонейронов у-мотонейроны не имеют непосредственного синаптического контакта с первичными афферентными волокнами. [10]
Возбуждение рецепторов мышечных веретен усиливается при увеличении механического сопротивления сокращению мышцы, в которой находятся веретена. Длина мышцы оказывается большей, чем при сокращении, преодолевающем меньшее сопротивление. Поэтому частота афферентных сигналов возрастает. Возбуждение рецепторов мышечных веретен вызывает рефлекс на растяжение - возрастает сила сокращений мышц, в которых находятся веретена. Сигналы от рецепторов веретен направляются в спинной мозг и усиливают возбуждение соответствующих а-мотонейронов. [11]
Страницы: 1