Экстремально галофильные архебактерия
Cтраница 1
 |
Представители группы не-фотосинтезирующих скользящих бактерий, не образующих плодовых тел. [1] |
Экстремально галофильные архебактерии, составляющие III подгруппу, представлены грамположительными или грамот-рицательными формами, аэробными или факультативно анаэробными хемоорганотрофами. Характерна потребность в высоких концентрациях NaCl. Некоторые виды содержат бактериородопсин и способны использовать энергию света для синтеза АТФ. В природе распространены в местах с высокой концентрацией соли: в соленых озерах, белковых продуктах, законсервированных с помощью соли, например в соленой рыбе. [2]
В девятом издании Определителя бактерий Берги экстремально галофильные архебактерии объединены в порядок Halobacteriales семейство Halobacteriaceae и включает 6 родов и более 20 видов, различающихся формой клеток ( палочки, кокки, квадраты), способностью к движению, отношением к кислотности среды, устойчивостью к NaCl и другими признаками. [3]
Вопрос о происхождении бесхлорофилльного фотосинтеза, обнаруженного у экстремально галофильных архебактерий, не ясен. [4]
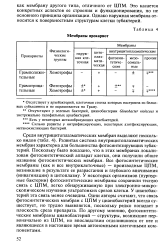 |
Мембраны прокариот. [5] |
Отсутствуют у зеленых бактерий, цианобактерии Gloeobacter violaceus и экстремально галофильных архебактерий. [6]
У прокариот известны три типа фотосинтеза: I - зависимый от бактериохлорофилла бескислородный фотосинтез, осуществляемый группами зеленых, пурпурных бактерий и гелиобактерий; II - зависимый от хлорофилла кислородный фотосинтез, свойственный цианобактериям и прохлорофитам; III - зависимый от бактериородопсина бескислородный фотосинтез, найденный у экстремально галофильных архебактерий. В основе фотосинтеза I и II типа лежит поглощение солнечной энергии различными пигментами, приводящее к разделению электрических зарядов, возникновению восстановителя с низким и окислителя с высоким окислительно-восстановительным потенциалом. Перенос электронов между этими двумя компонентами приводит к выделению свободной энергии. В фотосинтезе III типа окислительно-восстановительные переносчики отсутствуют. [7]
Такая же величина S характеризует и филогенетически далекие группы эубактерии. По молекулярной организации ветвь, включающая метанобразующие и экстремально галофильные архебактерии, ближе к эубактериям, а серозависимые архебактерии - к эукариотам. [8]
 |
Схема работы бакте. [9] |
Несмотря на кажущуюся простоту, очевидно, что бактериородопси-новая протонная помпа представляет собой сложную систему. Это означает, что поглощение кванта света должно приводить к возникновению напряженной конформации всего бактериородопсинового комплекса, служащей в дальнейшем источником энергии для переноса Н против электрохимического градиента. В организации такого переноса принимают участие ориентированные поперек мембраны а-спи-ральные тяжи и мембранные липиды, формирующие протонные каналы, природа и механизм действия которых пока не известны. Экстремально галофильные архебактерии содержат еще один ретинальбелковый комплекс - галородопсин, закачивающий на свету в клетки ионы хлора. Создающийся трансмембранный градиент СГ используется для синтеза АТФ. Одновременно этим обеспечивается также поддержание в цитоплазме высокой концентрации анионов, необходимой для уравновешивания высокой ионной силы внешней среды. [10]
Страницы: 1