Внутренняя митохондриальная мембрана
Cтраница 1
Внутренняя митохондриальная мембрана свободно проницаема для воды. [1]
 |
Транспортные системы внутренней мнтохондриальной мембраны, переносящие ADP и фосфат из цитозоля в матрикс, а новосинтезированный АТР-из матрикса в цито-золь. [2] |
Внутренняя митохондриальная мембрана непроницаема для наружного NADH, который находится в цитозоле. Каким же образом может NADH, образующийся в процессе гликолиза, который, как известно, протекает вне митохондрий, вновь окисляться с образованием NAD 1 молекулярным кислородом через дыхательную цепь. [3]

Во внутренней митохондриальной мембране локализуются переносчики электронов, составляющие дыхательную цепь, и ферменты, катализирующие синтез АТР из ADP и фосфата. [5]
Во внутренней митохондриальной мембране имеются транспортные системы для адениновых нуклеотидов, фосфата и ряда метаболитов. Перенос электронов тормозится при понижении концентрации ADP и ускоряется, когда концентрация ADP возрастает благодаря тем или иным клеточным процессам, сопровождающимся утилизацией ДТР. Скорости гликолиза, цикла лимонной кислоты и процесса окислительного фосфорилирова-ния согласованы между собой. Эта согласованность обеспечивается взаимосвязанными регуляторными механизмами, которые реагируют на величину отношения [ ATP ] / [ ADP ] [ Pf ] и на содержание некоторых наиболее важных метаболитов, отражающих энергетическое состояние клеток. [6]
Предотвращает движение адениннуклеотидов через внутреннюю митохондриальную мембрану путем вытеснения нуклеотидов из белка транслоказы, связанного с мембраной. Карбоксиатрактилозид также специфический ингибитор транспорта адениннуклеотида, но неконкурентный в отношении нуклеотидов. Растительные митохондрии относительно нечувствительны к атрактилозиду, но не к карбо-ксипроизводным. [7]
Предотвращает движение адениннуклеотидов через внутреннюю митохондриальную мембрану, препятствуя освобождению нуклеотидов из белка-носителя ( транс-локазы) со стороны матрикса. Бонгкрековая кис лота - транс-изомер по расположению двух карбоксильных гр. [8]
Сукцинатдегидрогеназа прочно связана с внутренней митохондриальной мембраной. Одна молекула этого фермента содержит один остаток ковалентно связанного FAD и два железо-серных центра; в одном из этих центров находятся два атома железа, а в другом-четыре. [9]
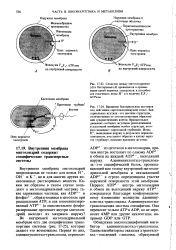
Адениннуклеотид-транслоказа-это специфический белок, пронизывающий всю толщу внутренней митохондриальной мембраны и связывающий ADP3 - в строго определенном участке наружной поверхности этой мембраны. Перенос ADP3 - внутрь митохондрии в обмен на выходящий наружу АТР - совершается благодаря конформацион-ному изменению молекулы аденинну-клеотид-транслоказы. [11]
Согласно этой гипотезе, функция переноса электронов, происходящего во внутренней митохондриальной мембране, заключается в том, чтобы откачивать ионы Н из матрикса митохондрии в наружную среду и таким путем создавать между двумя водными фазами, которые разделяет эта мембрана, градиент концентрации ионов Н с более кислым значением рН снаружи. Такой градиент, при котором концентрация ионов Н снаружи выше, чем внутри митохондрии, обладает потенциальной энергией ( разд. Хемиосмотическая гипотеза постулирует далее, что ионы Н, выведенные наружу за счет энергии переноса электронов, снова устремляются внутрь, в митохондриальный матрикс, через специальные каналы, или поры, для этих ионов в молекулах FgFj-АТРазы. В этом случае они перемещаются по градиенту концентрации и во время их перехода через молекулы АТРазы выделяется свободная энергия. Именно эта энергия и служит движущей силой для сопряженного синтеза АТР из ADP и фосфата. [12]

Малат покидает митохондрии при участии специальной дикарбоксилатной транспортной системы, находящейся во внутренней митохондриальной мембране ( разд. [14]
При 10 5 - 104 М разобщает окислительное фосфорилирование, переправляя поток протонов через внутреннюю митохондриальную мембрану; стимулирует митохондриальную АТРазу. [15]
Страницы: 1 2 3