Первичные анаэроб
Cтраница 1
Продукты первичных анаэробов не могут быть разложены далее с выигрышем энергии. Если условия складываются таким образом, Что развитие первичных анаэробов проходит очень быстро, вследствие обилия доступных веществ, прежде всего растворимых полисахаридов, то накопление продуктов останавливает развитие сообщества. Типичным примером служит компостирование растительных остатков и его бытовые варианты в виде квашения капусты. [1]
Главным продуктом первичных анаэробов и таких вторичных анаэробов, как гомоацетатные бактерии и I группа суль фатре-дукторов с неполным окислением, является ацетат. [2]
Бактероиды представляют группу типичных первичных анаэробов, сбраживающих углеводы в органические кислоты. Сюда относятся также Fusobacterium, Succinivibrio, Selenomonas, изогнутые клетки которого имеют мощный пучок жгутиков на вогнутой стороне. Бактероиды образуют совершенно самостоятельную филогенетическую ветвь. [3]
Таким образом, у ряда первичных анаэробов, получающих энергию в процессах брожения, сформировались короткие, связанные с мембраной электронтранспортные цепи, функционирование которых ведет к образованию протонного градиента, используемого для синтеза АТФ. Из-за отсутствия подходящего конечного акцептора электронов в анаэробных условиях выход энергии в такого типа процессах низкий. Однако принципиальные основы для создания энергетики нового типа сформированы. [4]

Обратимо функционирующие протонные АТФазы мы находим у первичных анаэробов, получающих энергию в процессе брожения. Обнаружено, что выделение во внешнюю среду молочной и уксусной кислот молочнокислыми бактериями и клостридиями приводит к созданию на ЦПМ протонного градиента. У стрептококков, осуществляющих гомоферментативное молочнокислое брожение, молочная кислота накапливается в клетке в виде аниона, для которого ЦПМ практически непроницаема. [6]
Аноксигенные фототрофы способны окислить практически все продукты первичных анаэробов ( Н2, ацетат, ЛЖК) и H2S как продукт сульфидогенов. При этом может образовываться и молекулярная сера, служащая субстратом для сероредуцирующих сульфидогенов с гораздо более широким кругом используемых веществ, чем у сульфатвосста-навливающих. Однако фотическая зона находится вне области действия гидролитиков, развивающихся в затененных донных осадках. Окислять метан фототрофные организмы не способны. [7]
Группы анаэробных гидролитиков и диссипотрофов объединяются в группировку первичных анаэробов, очень четко просматриваемую при анаэробных процессах как группировка бродилыциков, осуществляющая кислопгогенную или, иначе, водородную фазу. [8]
Субстрат роста - Н2 - служит характерным продуктом обмена первичных анаэробов. Для литоавтотрофного роста необходима автотрофная ассимиляция СО2, которая осуществляется через рибулозобисфосфатный цикл. Гидрогенотрофная автотрофия широко распространена среди бактерий, получивших название гидрогеномонады. [9]
Группировка вторичных анаэробов включает организмы с анаэробным окислительным обменом, обусловленным использованием несбраживаемых продуктов первичных анаэробов в качестве доноров электронов, и внешних неорганических акцепторов электронов в окислительно-восстановительных реакциях, приводящих к образованию полностью окисленного продукта разложения органического вещества - СО2 - и восстановленного неорганического соединения. [10]
Прямое окисление ЛЖК известно прежде всего у факультативно анаэробных денитрификаторов, для которых окисление всех продуктов первичных анаэробов представляет вполне банальную вещь и не привлекает специального внимания. Классическим примером такого организма явился Paracoccus denitrifi-cans, послуживший для К. Ван Ниля моделью для иллюстрации способности микробов осуществлять разные типы обмена ( Клюй-вер А. Действительно, паракокк хорошо растет аэробно с широким набором органических веществ. Он может расти хемолитоавтотрофно, как водородная бактерия, и относится к гидрогенотрофам. Хорошо выделяется на ацетате, как ацетотроф. [11]
Продукты первичных анаэробов не могут быть разложены далее с выигрышем энергии. Если условия складываются таким образом, Что развитие первичных анаэробов проходит очень быстро, вследствие обилия доступных веществ, прежде всего растворимых полисахаридов, то накопление продуктов останавливает развитие сообщества. Типичным примером служит компостирование растительных остатков и его бытовые варианты в виде квашения капусты. [12]
Таким образом, образование пары гидролитик - дисси-иотроф представляет правило. Для анаэробных целлюлозолитиков необходимо удаление продуктов брожения, так же как для многих первичных анаэробов. Поэтому целлюлозолитические организмы развиваются в тесных синтрофных отношениях с рядом сопутствующих форм. [13]
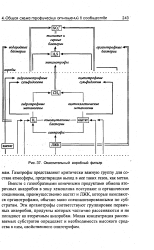 |
Окислительный аэробный фильтр. [14] |
Газотрофы представляют критически важную группу для состава атмосферы, предотвращая выход в нее таких газов, как метан. Вместе с газообразными конечными продуктами обмена вторичных анаэробов в зону хемоклина поступают и органические соединения, преимущественно ацетат и ЛЖК, которые окисляются органотрофами, обычно мало специализированными по субстратам. Эти органотрофы соответствуют группировке первичных анаэробов, продукты которых частично рассеиваются и не попадают ко вторичным анаэробам. Малая концентрация рассеиваемых субстратов определяет и необходимость высокого сродства к ним, свойственного олиготрофам. [15]
Страницы: 1 2