Триплет-триплетная аннигиляция
Cтраница 3
Эволюция во времени состояния электронно-дырочной пары, как показано Гроффом и др. [139], определяется спиновым гамильтонианом, который состоит из симметричного оператора сверхтонкого взаимодействия электрона и дырки и зеемановского взаимодействия. Эффекты, возникающие из-за анизотропии g - фактора и различия между g - факторами электронов и дырок, малы. Изменение заселенности триплетных состояний проявляется в модуляции замедленной флуоресценции, обусловленной триплет-триплетной аннигиляцией. На рис. 1.7.12 показана векторая диаграмма, иллюстрирующая синглет-триплетный интеркомбинационный переход пары радикалов в сильном магнитном поле. Электронные спины, обозначенные цифрами 1 и 2 для дырки и электрона, изменяют свои относительные фазы, так как каждый из них чувствует слегка различные локальные магнитные поля. Применение теории [140], подобной обсуждавшейся выше теории экситонной аннигиляции, для качественного анализа магнитной сверхтонкой модуляции сенсибилизированной красителем замедленной флуоресценции антрацена дает константу скорости диссоциации состояния электронно-дырочной пары & ] 1 107 с 1, которая оказывается очень низкой по сравнению с частотой прыжков дырок в антрацене. [31]
В заключение необходимо отметить, что формализм теории, развитый применительно к рекомбинации радикалов, можно легко обобщить и распространить на другие бимолекулярные процессы. Для этого в теорию достаточно ввести незначительные, непринципиальные модификации. Действительно, сравнение формального аппарата теории рекомбинации с теорией таких бимолекулярных процессов, как триплет-триплетная аннигиляция экситонов в кристаллах, тушение триплетных молеюлл парамагнитными добавками обнаруживает их общность. [32]
Только что описанный тип задержанной флуоресценции не обнаруживает такой же зависимости от температуры, как термически активированная задержанная флуоресценция Е - типа ( разд. Но более сильным отличительным признаком является зависимость сигнала флуоресценции от интенсивности поглощенного света. Эта зависимость линейна в случае задержанной флуоресценции Е - типа, но квадратична в случае процесса триплет-триплетной аннигиляции. [33]
Используя примесные кристаллы С10Н8 - C10Dg с концентрациями примесных молекул С 0Н8 от 0 1 до 20 %, они обнаружили, что ID - 1ХР, где х является функцией интенсивности возбуждения, интервала времени после возбуждения и концентрации примеси. При Т 1 7 К величины х лежат в интервале от 2 до - 30 и монотонно возрастают с уменьшением концентрации нафталина. Классическая кинетика справедлива для примесных кристаллов, когда концентрация примеси выше порога протекания ( в случае триплет-триплетной аннигиляции это соответствует С 10 % молярной доли); в этом случае / 1 и 7 не зависит от времени. Ранее было показано [10], что ограниченные кластером случайные блуждания имеют свойства, характерные для кинетики в условиях протекания. [34]

Похоже, что роль дефектов в генерации не очень значительна. Поскольку константа скорости взаимодействия триплетов с заполненными ловушками зависит от магнитного поля ( см. разд. В принципе, может проявляться и более тонкий эффект, связанный с возможным уменьшением в магнитном поле скорости триплет-триплетной аннигиляции. Вероятно, наблюдаемый эффект связан с влиянием внешнего магнитного поля на рекомбинацию электронов и дырок. В ионных красителях избыточные дырки нейтрализуют анионы, а свободные электроны приводят к появлению нейтральных хромофорных групп; образующиеся при этом продукты нейтрализации находятся в синглетном состоянии. С другой стороны, сами ионы являются дублетными состояниями. В отсутствие внешнего магнитного поля в результате рекомбинации могут образовываться пары ионов, находящиеся как в синглетном, так и в триплетном состоянии. Разница в энергиях этих состояний, по-видимому, невелика, поэтому конечное состояние ионной пары в значительной степени должно представлять собой смесь синглетных и триплетных состояний. Включение внешнего магнитного поля снимает вырождение между триплетными состояниями с разными проекциями спина и так же, как это происходит в случае взаимодействия трип-летного экситона с захваченным зарядом ( см. разд. [36]
Как и в случае свободной молекулы, переход из основного ( синглетного) состояния кристалла в триплетное экситонное состояние запрещен из-за правила отбора по спину. Как обсуждалось в разд. Поляризованный спектр поглощения перехода S0 - Tl в антрацене можно определить из измерений спектра возбуждения замедленной флуоресценции, которая возникает при триплет-триплетной аннигиляции. Для триплетных экситонов доминирует взаимодействие между ближайшими соседями; давыдовское расщепление приблизительно равно 81) 81, где / J - полная энергия взаимодействия между ближайшими неэквивалентными молекулами для нулевого вибронного возбужденного уровня, включающая также факторы Франка - Кондона. Дополнительный коэффициент 2 в теоретическом выражении WD для кристалла антрацена по сравнению с соответствующим выражением для линейной цепочки возникает из-за наличия четырех ближайших соседей в кристалле антрацена по сравнению с двумя для линейной цепочки. Молекулы, расположенные в антрацене перпендикулярно плоскости ab, взаимодействуют слабо по сравнению с молекулами, лежащими в самой плоскости, и их взаимодействием можно пренебречь. В этом смысле движение триплетных экситонов в антрацене почти двумерно ( см. разд. Экспериментальное значение WD 17 см 1 отвечает & 2 см 1, которое в три раза меньше соответствующего значения для 1 4 - DBN ( см. разд. [37]
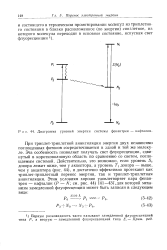 |
Диаграмма уровней энергии системы фенантрен - нафталин. [38] |
При триплет-триплетной аннигиляции энергия двух независимо поглощенных фотонов сосредоточивается в одной и той же молекуле. Эта особенность позволяет получать свет флуоресценции, сдвинутый в коротковолновую область по сравнению со светом, поглощаемым системой. Действительно, это возможно, если уровень S, донора лежит ниже, чем у акцептора, а уровень 7 донора - выше, чем у акцептора ( рис. 44), и достаточно эффективно протекают как триплет-триплетный перенос энергии, так и триплет-триплетная аннигиляция. [39]
Тем не менее связь еще может иметь место благодаря обменному члену, который зависит от перекрывания углеродных 2 / эя-орбиталей соседних молекул. Однако совсем недавно Найман и Робинсон [150] показали, что вследствие уменьшения с расстоянием эффективного заряда ядра до единицы и конфигурационного смешивания обменный интеграл может приобретать большое значение. Действительно, матричный элемент взаимодействия, связанного с триплетным состоянием, может быть величиной того же порядка, что и в случае низшего синглетно-го состояния, а именно порядка 100 см - г. Следствиями столь сильного взаимодействия, а также большого времени жизни триплетного состояния могут быть: а) большая возможность осуществления ( - 10П) актов передачи триплетного возбуждения за время жизни по отношению к излучению в чистых органических кристаллах по сравнению с меньшей возможностью в случае низшего возбужденного синглетного состояния ( только 104 - 106 актов); б) тушение фосфоресценции в чистых кристаллах при быстрой триплет-триплетной аннигиляции; в) миграция триплетного возбуждения от ловушки к ловушке; г) триплет-триплетная аннигиляция в случае кристаллов с примесями, приводящая к появлению замедленной флуоресценции; д) возможное усиление роли члена обменного взаимодействия в дополнение к члену кулоновского взаимодействия для синглетов; е) возможное влияние на биологические системы. [40]
Тем не менее связь еще может иметь место благодаря обменному члену, который зависит от перекрывания углеродных 2 / эя-орбиталей соседних молекул. Однако совсем недавно Найман и Робинсон [150] показали, что вследствие уменьшения с расстоянием эффективного заряда ядра до единицы и конфигурационного смешивания обменный интеграл может приобретать большое значение. Действительно, матричный элемент взаимодействия, связанного с триплетным состоянием, может быть величиной того же порядка, что и в случае низшего синглетно-го состояния, а именно порядка 100 см - г. Следствиями столь сильного взаимодействия, а также большого времени жизни триплетного состояния могут быть: а) большая возможность осуществления ( - 10П) актов передачи триплетного возбуждения за время жизни по отношению к излучению в чистых органических кристаллах по сравнению с меньшей возможностью в случае низшего возбужденного синглетного состояния ( только 104 - 106 актов); б) тушение фосфоресценции в чистых кристаллах при быстрой триплет-триплетной аннигиляции; в) миграция триплетного возбуждения от ловушки к ловушке; г) триплет-триплетная аннигиляция в случае кристаллов с примесями, приводящая к появлению замедленной флуоресценции; д) возможное усиление роли члена обменного взаимодействия в дополнение к члену кулоновского взаимодействия для синглетов; е) возможное влияние на биологические системы. [41]
Триплет-триплетная аннигиляция при импульсном возбуждении наблюдается очень часто, особенно при больших концентрациях триплетных молекул. Частично этот процесс связан с переводом одной из триплетных молекул в синглетнр-возбуж-денное, а второй - в основное состояние. Константа скорости триплет-триплетной аннигиляции является диффузионной и составляет 109 - f - 1010 М - - с 1 для обычных невязких растворителей. Энергия активации триплет-триплетной аннигиляции совпадает с энергией активации диффузии. Таким образом, лимитирующей стадией триплет-триплетной аннигиляции является диффузия триплетных молекул. [42]
Механизм этой аннигиляции состоит в следующем. При столкновении двух триплетных экситонов возможна передача энергии, в результате чего одна из молекул может оказаться в синглетном возбужденном состоянии. Наблюдаемой величиной, которая свидетельствует о триплет-триплетной аннигиляции, может служить замедленная флуоресценция синглетных возбужденных молекул. [43]
Формула ( 417) эквивалентна уравнению, впервые полученному Бирксом [279] для случая замедленной флуоресценции, возбуждаемой светом. Константы Кз и К2 [ см. уравнение ( 409) ], рассчитанные Паркером и Хатчардом для пирена в этаноле, эквивалентны константам К. Паркер и Шорт [280] установили, что при электрохемилюминесценции 9 10-диметилантрацена в диметилформамиде испускание мономера и димера подчиняется уравнению ( 419), причем обе константы увеличиваются при снижении температуры и напряжения. Значение К при низкой температуре и низком напряжении оказалось гораздо больше, чем значение Kz [ см. уравнение ( 409) ], найденное Паркером и Джойс [179] для замедленной флуоресценции, возбуждаемой при тех же условиях светом; это указывало, что триплет-триплетная аннигиляция не была главным источником электрохемилюминесценции. [44]
Триплет-триплетная аннигиляция при импульсном возбуждении наблюдается очень часто, особенно при больших концентрациях триплетных молекул. Частично этот процесс связан с переводом одной из триплетных молекул в синглетнр-возбуж-денное, а второй - в основное состояние. Константа скорости триплет-триплетной аннигиляции является диффузионной и составляет 109 - f - 1010 М - - с 1 для обычных невязких растворителей. Энергия активации триплет-триплетной аннигиляции совпадает с энергией активации диффузии. Таким образом, лимитирующей стадией триплет-триплетной аннигиляции является диффузия триплетных молекул. [45]
Страницы: 1 2 3