Гормон-рецепторный комплекс
Cтраница 1
Гормон-рецепторный комплекс проникает в ядро и связывается с хроматином в области андрогенчувствительного элемента ДНК. [1]
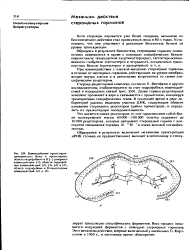 |
Взаимодействие прсгестерон. [2] |
Далее гормон-рецепторный комплекс проникает в ядро и связывается с хроматином, инициируя транскрипцию специфических генов. [3]
Гормон, взаимодействуя с К, образует гормон-рецепторный комплекс НогК, к-рый, реагируя с О-ОЙР ( см. схему), вызывает каскад р-ций. Непосредственно катализирует превращение АТФ в цАМФ комплекс С8 - [ С-СТР ], к-рый в клетке быстро распадается на С8 и С-СОР. [4]
При взаимодействии стероидного гормона с рецептором образуется гормон-рецепторный комплекс, который мигрирует далее в клеточное ядро. Такой комплекс имеет большее сродство к ДНК, чем несвязанный рецептор. Наличие такого эффекта показано для эстрадиола, но, очевидно, он возможен и для других гормонов. [5]
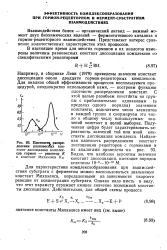 |
Плотности, распределения равновесных констант диссоциация комплексов гормон - рецептор К. [6] |
Например, в сборнике Леви ( 1979) приведены значения констант диссоциации около двадцати гормон-рецепторных комплексов. Для анализа общей эффективности процессов воспользуемся процедурой, неоднократно используе. [7]
Так же как и другие стероиды, эстрогены проникают в цитоплазму и далее в составе гормон-рецепторного комплекса в ядро клеток. Эстрогены не влияют непосредственно на РНК-полимеразу, но резко стимулируют процессы инициации транскрипции. Усиление синтеза специфических белков в репродуктивных органах инициирует ряд зависимых процессов на клеточном и органном уровнях. Так, эстрогены контролируют процессы овуляции за счет индукции секреции лютеинизирующего гормона по принципу обратной связи. Прогестерон обеспечивает эффективность имплантации оплодотворенной яйцеклетки в матке, а также стабилизирует мембранный потенциал миометрия, что способствует сохранению беременности. [8]
 |
Участок взаимодействия гормон-рецепторного комплекса с ДНК. [9] |
Активированный гормон-рецепторный комплекс связывается с ядерными мембранами и перемещается в ядро клетки. [10]
Кор-тикостероиды, а также эстрогены и ан-дрогены проходят через плазматические мембраны соответствующих клеток-мишеней и связываются с рецепторами в цитозоле. Далее гормон-рецепторный комплекс воздействует на ядро, вызывая экспрессию определенных генов. [11]
После освобождения гормон-рецепторного комплекса и его дефосфорилирования ядерными фосфопротеинфосфатазами происходит диссоциация комплекса на гормон и рецептор, последний перемещается в цитоплазму, ассоциируется с белками теплового шока и включается в следующий цикл передачи гормонального сигнала. Этот процесс называется рециклизацией гормонального рецептора. В некоторых случаях рециклизация не происходит, так как рецептор после поступления из ядра в цитоплазму подвергается протеолитическому расщеплению. [12]
В репродуктивных тканях андрогены отвечают за их дифференцировку и функционирование. Образовавшийся в семенниках тестостерон и его активный метаболит ДГТ проникают в клетки-мишени методом простой или облегченной диффузии и взаимодействуют с одним и тем же белковым рецептором. Образовавшиеся гормон-рецепторные комплексы перемещаются в ядро, связываются с хроматином и стимулируют процессы синтеза белка ( гл. В репродуктивных органах эти процессы реализуются в половой дифференцировке, основные этапы которой представляют собой: хромосомы-гонады-фенотип. Кроме того, андрогены стимулируют сперматогенез, половое созревание и по принципу обратной связи контролируют секрецию гонадотропинов. Помимо влияния на функционирование репродуктивной системы, андрогены участвуют в контроле клеточного метаболизма многих других тканей и органов. Независимо от типа ткани андрогены проявляют анаболические эффекты, связанные со стимуляцией процессов транскрипции и увеличения скорости синтеза белка. Более всего андрогенных клеток-мишеней находится в скелетных мышцах, причем под действием гормонов происходит резкое увеличение мышечных белков и наращивание мышечной массы. Стимуляция белок-синтетических процессов под действием андрогенов отмечена в почках, сердечной мышце, костной ткани. Андрогены образуются не только в семенниках, но и в яичниках. Их роль в организме женщин или самок животных заключается в формировании поведенческих реакций, а также в контроле за синтезом белка в репродуктивных органах. [13]
Известно, что АКТГ, стимулируя аденилатциклазу коркового слоя надпочечников, вызывает значительное увеличение содержания цАМФ, который в свою очередь по протеинкиназному механизму обеспечивает синтез короткоживущего лабильного белка. Последний участвует в процессе превращения холестерина в прегненолон [ Gar-геп L. Можно предположить, что при облучении над-лочечников in vitro нарушается взаимодействие гормон-рецепторного комплекса с аденилатциклазой, угнетается синтез цАМФ и корти-жостероидов. Однако in vivo не наблюдается снижение продукции гормонов коры надпочечников, более того, обнаружены фазы подъема в плазме крови уровня кортикостероидов в различные периоды лучевой болезни [ Мороз Б. Б., Ромашко О. О., 1978; Лицкевич Л. А. ж др., 1978; Groza P. [14]
Одной из основных функций инсулина является регуляция транспорта глюкозы, аминокислот, ионов и других метаболитов в клетки печени, почек, жировой ткани и других органов. Механизм действия этого гормона отличается от такового для других пептидных гормонов и является уникальным в регуляции метаболических процессов. Инсулиновый рецептор представляет собой тетрамер, состоящий из двух а - и двух р-субъеди-ниц, одна из которых обладает тирозинкиназной активностью. Инсулин при взаимодействии с а-субъединицами, расположенными на поверхности цито-плазматической мембраны, образует гормон-рецепторный комплекс. Кон-формационные изменения тетрамера приводят к активации трансмембранной ( 3-субъединицы рецептора, обладающей тирозинкиназной активностью. Активная тирозинкиназа способна как к аутофосфорилированию, так и к локальному фосфорилированию близлежащих мембранных белков. В результате фосфорилирования образуются мембранные каналы, через которые глюкоза и другие метаболиты проникают в клетки. Через определенный временной интервал происходит интернализация гормон-рецепторного комплекса вовнутрь клетки, где он диссоциирует на свободные рецептор и гормон. Рецептор поступает в аппарат Гольджи, где ремонтируется, а затем перемещается на внешнюю мембрану для дальнейшего функционирования. Этот процесс называется рециклизацией инсулинового рецептора. Свободный инсулин под действием тканевой инсулиназы распадается на семь фракций, пять из которых обладают биологической активностью. Так, они способны активировать бБ - белки рибосом, обладающие протеинкиназной активностью. В результате стимулируется инициация трансляции и биосинтез белка. [15]
Страницы: 1 2