Гомологичная рекомбинация
Cтраница 4
Системы репликации и репарации ДНК хоть и очень редко, но все же ошибаются. В результате в ДНК может включиться неправильный, т.е. не комплементарный матрице, нуклеотид. Другой источник неспаренных нуклеотидов - гомологичная рекомбинация, в ходе которой образуются гетеродуплексы, состоящие из двух комплементарных цепей, исходно принадлежавших разным молекулам ДНК ( см. гл. Такие нарушения структуры ДНК репари-руются, по крайней мере у бактерий и низших эукариот. [46]
Рекомбинация у эукариотических клеток была выявлена генетическими методами, а в отдельных случаях и путем наблюдения форм хромосом. Этот процесс происходит при созревании половых клеток, на первой фазе которого две пары хромосом, образовавшиеся в результате предшествующей репликации, вместо того чтобы разойтись по двум дочерним клеткам, как это имеет место при обычном клеточном делении - митозе, предварительно объединяются в единую структуру некоторыми гомологичными сегментами. Это создает благоприятные условия для гомологичной рекомбинации, которая у эукариот, в первую очередь у дрозофилы, была открыта задолго до выяснения рекомбинации у бактерий и получила название кросситовера. Рекомбинация сама по себе не создает новых генов, однако в результате нее возникают новые комбинации признаков, которые могут оказаться весьма существенными как при естественном отборе, так и в селекционных работах. [47]
Клонирующий вектор несет участок Т - ДНК, содержащий клонированный ген. После введения в клетку Agrobacterium он подвергается гомологичной рекомбинации с резидентной разоруженной ( неонко-генной) Ti-плазмидой с образованием одной плазмиды, несущей генетическую информацию, необходимую для переноса генетически измененной области Т - ДНК в растительную клетку. [48]
Общая, или гомологичная, рекомбинация характерна для всех живых организмов от вирусов и бактерий до многоклеточных эука-риот. ДНК - Так, к сбщей рекомбинации относятся обмены между гомологичными хромосомами в мейозе у эукариот и реком-бинацнонная инициация репликации ДНК бактериофага Т4 ( см. гл. Чтобы подчеркнуть важность этого свойства, достаточно сказать, что при гомологичной рекомбинации между двумя сходными генами, кодирующими белок, оба рекомбинантных продукта оказываются не нарушенными, не происходит, например, сдвига рамки считывания. Другими словами, при гомологичной рекомбинации каким-то образом обеспечивается взаимное узнавание одинаковых ( или очень сходных по последовательности) участков рекомбинирующих молекул. [49]
Общая, или гомологичная, рекомбинация характерна для всех живых организмов от вирусов и бактерий до многоклеточных эука-риот. ДНК - Так, к сбщей рекомбинации относятся обмены между гомологичными хромосомами в мейозе у эукариот и реком-бинационная инициация репликации ДНК бактериофага Т4 ( см. гл. Чтобы подчеркнуть важность этого свойства, достаточно сказать, что при гомологичной рекомбинации между двумя сходными генами, кодирующими белок, оба рекомбинантных продукта оказываются не нарушенными, не происходит, например, сдвига рамки считывания. Другими словами, при гомологичной рекомбинации каким-то образом обеспечивается взаимное узнавание одинаковых ( или очень сходных по последовательности) участков рекомбинирующих молекул. [50]

Из библиотеки кДНК, которая была получена на основе мРНК, выделенной из биссаловой железы, была изолирована кДНК 130 кДа - пред-шественника адгезивного белка. И адгезивный белок, и его кДНК обладают весьма необычными свойствами, затрудняющими клонирование и экспрессию соответствующих генов и получение функционального адгезивного белка. Во-первых, кДНК содержит большое число повторов, что повышает частоту гомологичной рекомбинации и вероятность утраты части клонированной последовательности. Во-вторых, поскольку примерно 70 % всех аминокислот белка приходится на долю пролина, лизина и тирозина, вряд ли его удастся получить в большом количестве вследствие ограниченности внутриклеточного пула аминоацил-т РНК. [52]
Во втором случае используют коинтегративную векторную систему. Коинтегративный вектор и неонкогенная Ti-плазмида-помощник содержат гомологичные последовательности, которые образуют сайт для гомологичной рекомбинации in vivo. Обычно эти последовательности расположены в Т - ДНК. После рекомбинации клонирующий вектор становится частью неонкогенной Ti-плазми-ды, которая содержит v / r - гены, необходимые для переноса Т - ДНК в растительную хозяйскую клетку. [53]
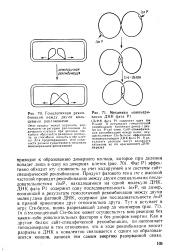 |
Гомологичная рекомбинация между двумя кольцевыми репликонами.| Механизм мономери. [54] |
Фаг Р1 эффективно обходит эту сложность за счет кодируемой им системы сайт-специфической рекомбинации. Продукт фагового гена ere с высокой частотой проводит рекомбинацию между двумя специальными последовательностями / охР, находящимися на одной молекуле ДНК. ДНК фага Р1 содержит одну последовательность loxP, но димер, возникший в результате гомологичной рекомбинации между двумя молекулами фаговой ДНК, содержит две последовательности loxP в прямой ориентации друг относительно друга. In vitro очищенный Сге-белок может осуществлять всю реакцию без каких-либо дополнительных факторов и без доноров энергии. Как и другие белки сайт-специфической рекомбинации, он напоминает топоизомеразы в том отношении, что в ходе рекомбинации вносит разрывы в ДНК и ковалентно связывается с одним из образовавшихся концов, запасая тем самым энергию разорванной связи. [55]
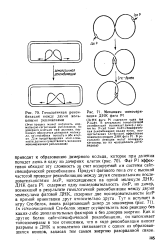
Фаг Р1 эффективно обходит эту сложность за счет кодируемой им системы сайт-специфической рекомбинации. Продукт фагового гена ere с высокой частотой проводит рекомбинацию между двумя специальными последовательностями loxP, находящимися на одной молекуле ДНК. ДНК фага Р1 содержит одну последовательность loxP, но димер, возникший в результате гомологичной рекомбинации между двумя молекулами фаговой ДНК, содержит две последовательности / о. Сге-белок может осуществлять всю реакцию без каких-либо дополнительных факторов и без доноров энергии Как и другие белки сайт-специфической рекомбинации, он напоминает топоизомеразы в том отношении, что в ходе рекомбинации вносит разрывы в ДНК и ковалентно связывается с одним из образовавшихся концов, запасая тем самым энергию разорванной связи. [57]
Общая, или гомологичная, рекомбинация характерна для всех живых организмов от вирусов и бактерий до многоклеточных эука-риот. ДНК - Так, к сбщей рекомбинации относятся обмены между гомологичными хромосомами в мейозе у эукариот и реком-бинацнонная инициация репликации ДНК бактериофага Т4 ( см. гл. Чтобы подчеркнуть важность этого свойства, достаточно сказать, что при гомологичной рекомбинации между двумя сходными генами, кодирующими белок, оба рекомбинантных продукта оказываются не нарушенными, не происходит, например, сдвига рамки считывания. Другими словами, при гомологичной рекомбинации каким-то образом обеспечивается взаимное узнавание одинаковых ( или очень сходных по последовательности) участков рекомбинирующих молекул. [58]
Страницы: 1 2 3 4