Инициация
Cтраница 4
Время инициации ветви является величиной, не зависящей от времени выполнения программы ветви, но оказывающей влияние на целесообразный объем ( минимальное число команд) программы ветви. В таких случаях параллельные ветви должны быть принудительно сведены в одну последовательную ветвь. [46]
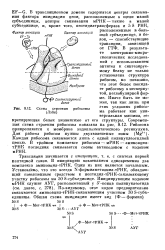 |
Схема строения рибосомы ( Лэйк. [47] |
В инициацию вовлекаются одновременно два комплекса аминоацил-т РНК. [48]
В инициацию должны вовлекаться одновременно два комплекса аминоацил-т РНК. [49]
Что касается инициации на концах одноцепочечной ДНК-мат-РИЦЫ, то тут известны два механизма. [50]
Что касается инициации на внутренних участках двухнитевой матрицы, то здесь также нужно различать два основных способа. Однако синтез затравки возможен только в том случае, если матрица соответствующим образом подготовлена. Подготовка включает взаимодействие между вирус-специфическими белками, регулирующими инициацию раунда репликации, и специфическими участками инициации репликации ori ( от англ, origin - начало) в молекуле ДНК - Например, с участком ori в ДНК фага Я первично взаимодействует фагоспецифический белок - О, с белком О взаимодействует другой фагоспецифический полипептид - белок Р, который Ш свою очередь образует комплекс с одной из клеточных хеликаз - продуктом гена dna В. [51]
Что касается инициации на концах одноцепочечной ДНК-матрицы, то тут известны два механизма. [52]
Что касается инициации на внутренних участках двухнитевой матрицы, то здесь также нужно различать два основных способа. Однако синтез затравки возможен только в том случае, если матрица соответствующим образом подготовлена. Подготовка включает взаимодействие между вирус-специфическими белками, регулирующими инициацию раунда репликации, и специфическими участками инициации репликации ori ( от англ, origin - начало) в молекуле ДНК - Например, с участком ori в ДНК фага h первично взаимодействует фагоспецифический белок - О, с белком О взаимодействует другой фагоспецифический полипептид - белок Р, который В свою очередь образует комплекс с одной из клеточных хеликаз - продуктом гена dna В. [53]
Что касается инициации на концах одноцепочечной ДНК-мат-РИЦЫ, то тут известны два механизма. [54]
Что касается инициации на внутренних участках двухнитевой матрицы, то здесь также нужно различать два основных способа. Однако синтез затравки возможен только в том случае, если матрица соответствующим образом подготовлена. Подготовка включает взаимодействие между вирус-специфическими белками, регулирующими инициацию раунда репликации, и специфическими участками инициации репликации ori ( от англ, origin - начало) в молекуле ДНК - Например, с участком ori в ДНК фага X первично взаимодействует фагоспецифический белок - О, с белком О взаимодействует другой фагоспецифический полипептид - белок Р, который свою очередь образует комплекс с одной из клеточных хеликаз - продуктом гена ana В. [55]
Порог для инициации микронеустойчивости в солнечной короне примерно равен 80 А / м2 и, похоже, единственное место, где он превышен - это интенсивные токовые концентрации, такие как токовые слои. Хейвартс и Куперус вывели различные режимы поведения токового слоя, зависящие от его длины и ширины. Они ввели короткий гидродинамический масштаб времени т l / cs 10 - 2 / / Т1 / 2 с, длинный гидродинамический масштаб времени TL L / cs 10 - 2L / T1 / 2 с, характерное время диффузии или джоулевого нагрева т / 2 / rj 10 - 9 / 2Т3 / 2 с, характерное время излучения ту p / ( n Q) 5 10 - 52 / / T2JB 02Q - 1 с и характерное время проводимости т р / 2 / ( Тх) с, при выводе которого предполагался баланс давлений поперек слоя. [56]
Гликогенсинтетаза для инициации реакции требует затравки полиглюкозы, имеющей не менее четырех остатков. Амило - ( 1 4 -у 1 6) - трансглюкозилаза осуществляет ветвление гликогена путем образования х - ( 1 - 6) - связей. [57]
Так или иначе внутренняя инициация происходит без разрыва родительских цепей ДНК. Первая стадия этого процесса заканчивается образованием двух дочерних дуплексов. Эти дуплексы имеют очень важную особенность - 5 -концы их вновь синтезированных цепей недостроены; другими словами, не произошло копирования З - концов родительских цепей. Такая ситуация неизбежно возникает при внутренней инициации на линейной ДНК-матрице. Причина нехватки понятна и является следствием двух универсальных свойств ДНК-полиме-раз: потребности в затравке и способности синтезировать цепочку - ДНК только в одном направлении - от 5 -конца к З - концу. [58]
Так или иначе внутренняя инициация происходит без разрыва родительских цепей ДНК. Первая стадия этого лроцесса заканчивается образованием двух дочерних дуплексов. Эти дуплексы имеют очень важную особенность - 5 -концы их вновь синтезированных цепей недостроены; другими словами, не произошло копирования З - концов родительских цепей. Такая ситуация неизбежно возникает при внутренней инициации на линейной ДНК-матрице. Причина нехватки понятна и является следствием двух универсальных свойств ДНК-полиме-раз: потребности в затравке и способности синтезировать цепочку - ДНК только в одном направлении - от 5 -конца к 3 -концу. [59]
Страницы: 1 2 3 4