Первичный акцептор
Cтраница 3
В приведенной схеме выражение молекул кофермента обозначает комплексную группу, обратимо присоединяющую пару электронов или водород. Обычно первичным акцептором водорода бывает ди - или трифосфопиридиннуклеотид или фла-вопротеин. Реакция ( 1) в энергетическом отношении несущественна, более того, почти вся свободная энергия дыхания возникает в реакции ( 2), сопряженной с синтезом АТФ. Следовательно, восстановленные коферменты являются первоначальными продуктами биологического окисления, причем они окисляются не полностью, но каждая молекула отдает 2 атома водорода. [31]
Первоначальный этап фотосинтеза в хлоропластах заключается в отрыве электронов, за счет энергии светового кванта, от комплекса молекул хлорофилла а, входящих в реакционные Фотосинтезирующие центры РК. Оторвавшиеся электроны переносятся на первичные акцепторы Аг, природа которых в различных фотосинтезирующих системах еще не вполне установлена. [32]
В основе фотосинтетического преобразования солнечной энергии лежит фотоиндуцируемый перенос электрона. В этом процессе роль донора и первичного акцептора электрона играют восстановленные тетрапир-рольные системы - хлорофилл и феофитин, имеющие макроциклическую конъюгированную систему л-электронных облаков. Функцию вторичного акцептора электрона выполняет молекула хинона. Порфирин-хиноновые диады широко используются в исследованиях фотоиндуцируемого переноса электрона в качестве биомиметических донорно-акцепторных модельных систем. Введение в их состав аминокислотных заместителей может имитировать присутствие аминокислотных остатков белкового окружения хромофоров в природных реакционных центрах. Для изучения воздействия присоединенной аминокислоты на фотохимические свойства пигмента были также получены ди - и монопроизводные дейтеропорфирина IX с теми же аминокислотами. Также были синтезированы аминокислотные производные замещенного тетрафенилпорфирина с целью выяснения характера влияния типа замещения порфиринового макроцикла и места присоединения аминокислоты на энергетику системы. Методами электронной и флуоресцентной спектроскопии изучены фотохимические свойства полученных модельных систем. Введение аминокислотного заместителя практически не влияет на положения максимумов полос поглощения и флуоресценции, однако, приводит к увеличению их интенсивности. Этот эффект зависит от места присоединения и природы аминокислоты, а также от конформации молекулы. [33]
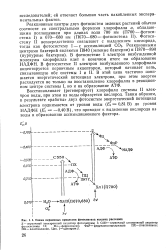 |
Схема первичных процессов фотосинтеза высших растений. [34] |
В фотосистеме I электрон возбужденной молекулы хлорофилла идет в конечном итоге на образование НАДФН. В фотосистеме II электрон возбужденного хлорофилла акцептируется первичным акцептором, который начинает цепь, связывающую обе системы I и II. В этой цепи частично понижается энергетический потенциал электрона, при этом энергия расходуется не только на восстановление хлорофилла в реакционном центре системы I, но и на образование АТФ. [35]
Исключительно важный процесс аккумулирования солнечной энергии путем образования органического вещества из СО2 и Н2О, называемый фотосинтезом, в последние годы в главных своих чертах был расшифрован ( Кэлвин и др.), что нужно считать одним из крупнейших достижений химии. При этом была выявлена очень важная роль фосфатов Сахаров: не только первичным акцептором углекислоты ( веществом, присоединяющим СО2) оказался фосфат моносахарида, но и круговой процесс ( так называемый фотосинтетический углеродный цикл), вновь образующий этот акцептор, в основном оказался сложным процессом превращений фосфорнокислых эфиров моносахаридов. [36]
В то время как основная масса фотосинтетических пигментов способна только поглощать энергию света и передавать ее соседним молекулам, небольшая часть молекул хлорофилла участвует в осуществлении фотохимической реакции, т.е. преобразовании электромагнитной энергии в химическую. Процесс этот происходит в реакционных центрах, состоящих из первичного донора электронов, первичного акцептора и одного или более вторичных акцепторов электронов. Кроме того, в составе реакционных центров обнаружены молекулы каротиноидов и полипептидов. [37]
Сахароза может служить донором не только остатков глюкозы, но и остатков фруктозы. Многие бактерии вырабатывают фермент, катализирующий образование ле ванов из сахарозы75 76; первичным акцептором, начинающим полисахаридную цепь, здесь также может быть сахароза. Наличие глюкозы в качестве концевого моносахарида у фруктанов высших растений позволяет предполагать аналогичный путь синтеза и для этих соединений. Точно так же содержание значительного количества маннита на концах полисахаридных цепей ламинарина ( см. стр. [38]
Представляют интерес работы, в которых проведены оценки времен электронных переходов п фотосинтезирующих системах пурпурных бактерий. При поглощении света происходит разделение зарядов в фотосинтеэирующем центре / 870t образуется окисленный донор D и возбужденный электрон переходит на первичный акцептор электронов 41 ( по-видимому, за время, измеряемое пикосекундами. Окисленный донор D получает электрон от цитохрома с ( см. рис. 45) в среднем в течение нескольких миллисекунд. [39]
Ренгерсом [225] проведено наиболее полное исследование фотосистемы ФСП. Реакционный центр ориентирован в мембране тилакоида так, что первичный донор электронов СЫац расположен с внутренней стороны мембраны, а первичный акцептор электронов Xj2o - вблизи внешней поверхности мембраны. [40]
Удаление пластохинона из хлоропластов путем экстракции приводит к инактивации фотосистемы II ( разд. Хотя эта инактивация с последующим восстановлением активности наглядно свидетельствует о роли пластохинона в транспорте электронов, такую относительно неспецифическую реактивацию, по-видимому, трудно согласовать с предполагаемой функцией пластохинона как первичного акцептора электрона во второй фотореакции. [41]
В книге излагаются современные представления о механизме рвичного акта преобразования энергии света в энергию разделенных рядов в реакционных центрах фотосинтеза, представляющих собой юрофилл-белковые комплексы мембран. Экспериментально пока-но, что формирование волновых пакетов при фемтосекундном збуждении первичного донора электрона, димера бактериохло-крилпа Р, приводится к ситуации, когда наблюдается отчетливая висимость переноса электрона от движения ядерной подсистемы оль координаты переноса электрона с Р на первичный акцептор [ ектрона, бактериохлорофилл ВА. Полученные результаты и их еретическое обоснование проливают свет на общие закономерности зделения зарядов как в биологических, так и в химических системах. [42]
Согласно современным представлениям наиболее вероятная схема переноса электронов по дыхательной цепи внутренних мембран митохондрий может быть описана следующим образом. От питательных веществ ( пируват, сукцинат, малат и др.) - субстратов ( S 2Н) - ферменты - пиридинзависимые дегидрогеназы - переносят два атома водорода ( два электрона) к молекулам НАД, которые выступают как первичные акцепторы электронов. При этом свободные ионы Н переходят во внутреннюю область митохондрий ( матрикс), где они вместе с ионами гидроксила ОН - образуют молекулы воды. [43]
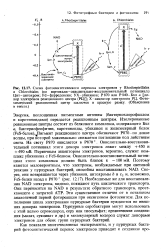
Энергия, поглощенная пигментами антенны ( бактериохлорофиллом и каротиноидами) передается реакционным центрам. Пигмент реакционного центра обозначают Р870 - по длине волны, при которой максимально снижается поглощение под действием света. Первичным акцептором электронов, вероятно, служит комплекс убихинона с FeS-белком. Поэтому кажется маловероятным, что электроны, возбуждаемые при световой реакции у пурпурных бактерий, способны восстанавливать NAD. Необходимые же для восстановления NAD электроны, видимо, покидают путь циклического транспорта. Они переносятся на NAD в результате обратного транспорта, протекающего с затратой АТР. [45]
Страницы: 1 2 3 4