Мутирование
Cтраница 4
Авторы [17] предполагают, что соседние основания слева и справа могут иметь большое значение для стабилизации неканонической пары, возникапцей в ходе репликации. Похожие закономерности выявлены при анализе спонтанных мутаций, возникающих при трансфекции вектора pZI89 в клетки обезьяны. Консенсус имеет вид: УС, где нуклеотид С - позиция мутирования. [46]
У человека также, безусловно, возникают спонтанные мутации, и делались попытки определить при помощи различных методов частоту мутирования разных генов. Простейший из этих методов основан на том, что в редких случаях у одного из детей может проявиться доминантный признак, отсутствующий у обоих родителей. Это должно свидетельствовать о возникновении признака вследствие новой мутации. Проведенное в Дании изучение наследования так называемой хондроди-строфии показало, что примерно одна гамета на 24 000 несет доминантную мутацию, обусловливающую это заболевание. Для 5 других наследственных заболеваний были найдены как более высокие, так и более низкие частоты мутирования. Средняя частота мутаций у человека, видимо, по крайней мере так же высока, как у дрозофилы, но данные, на которых основан этот вывод, еще недостаточно достоверны. [47]
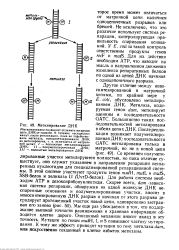 |
Метилирование ДНК. [48] |
ДНК - Метилаза, кодируемая геном dam, метилирует аденины в последовательности GATC. Со временем полуметилированные участки метилируются полностью, но пока отличие существует, оно служит указанием о направлении репарации неспаренных нуклеотидов для специализированной репарационной системы. Для работы системы необходим АТР и дезоксирибонуклеотиды. Мы еще раз убеждаемся в том, что поддержание высокой точности размножения и хранения информации обходится клетке дорого. Описанный механизм вносит вклад в эту точность. Мутации по генам mutH, L и S повышают частоту мутирования. К тому же эффекту приводят мутации по гену метил азы dam, или искусственно созданный в клетке избыток метил азы. [49]
Исходя из наблюдаемой скорости появления точковых мутаций ( одна мутация на 106 удвоений гена), мы можем подсчитать, что одна мутация приходится на 109 репликаций единичного нуклеотида. Точ-ковые мутации имеют тенденцию к обратному мутированию, причем обратные мутации часто происходят с такой же скоростью, как и прямые. Это значит, что в одной из 109 обратных мутаций будет мутировать тот же самый нуклеотид, в результате чего ген вернется к исходному виду. Это явление легко можно объяснить. Например, если Т будет замещен на С, поскольку С образует минорный таутомер и спаривается с А, то мутация приведет к тому, что в двойной спирали ДНК-потомков появится пара GC. При репликации этой пары существует хотя и малая, но определенная вероятность того, что С в цепи материнской ДНК вновь образует минорную таутомерную структуру и образует пару с А, а не с G, что в свою очередь приведет к обратному мутированию. [50]
 |
Метилирование ДНК. [51] |
ДНК - Метилаза, кодируемая геном dam, метилирует аденины в последовательности GATC. Большинство таких последовательностей метилированы в обеих цепях ДН К. Со временем полуметилированные участки метилируются полностью, но пока отличие существует, оно служит указанием о направлении репарации неспаренных нуклеотидов для специализированной репарационной системы. Для работы системы необходим АТР и дезоксирибонуклеотиды. Мы еще раз убеждаемся в том, что поддержание высокой точности размножения и хранения информации обходится клетке дорого. Описанный механизм вносит вклад в эту точность. Мутации по генам muffl, L и S повышают частоту мутирования. К тому же эффекту приводят мутации по гену метил азы dam, или искусственно созданный в клетке избыток метил азы. [52]
 |
Метилирование ДНК. [53] |
ДНК - Метилаза, кодируемая геном dam, метилирует аденины в последовательности GATC. Большинство таких последовательностей метилированы в обеих цепях ДНК. Со временем полуметилированные участки метилируются полностью, но пока отличие существует, оно служит указанием о направлении репарации неспаренных нуклеотидов для специализированной репарационной системы. Для работы системы необходим АТР и дезоксирибонуклеотиды. Мы еще раз убеждаемся в том, что поддержание высокой точности размножения и хранения информации обходится клетке дорого. Описанный механизм вносит вклад в эту точность. Мутации по генам mutH, L и S повышают частоту мутирования. К тому же эффекту приводят мутации по гену метилазы darn, или искусственно созданный в клетке избыток метилазы. [54]
Недавно было показано, что в генах V-области и поблизости от них происходят соматические мутации; вероятно, это увеличивает число разных антител по крайней мере в 10 - 100 раз. Механизмы, благодаря которым мутации возникают именно здесь, неизвестны. Однако было обнаружено, что мутации встречаются гораздо чаще в антителах IgG и IgA, чем в кодируемых тем же Ун-геном антителах IgM. Это, пожалуй, неудивительно, так как молекулы IgM вырабатываются на ранних стадиях иммунного ответа, a IgG и IgA появляются относительно поздно. Поэтому В-клетки, переключившиеся с IgM на IgG или IgA ( см. разд. С другой стороны, может существовать механизм, повышающий частоту мутирования V-генов после того, как произойдет переключение с IgM на иммуноглобулины других классов. [55]
ДНК-пслимеразы проверяют комплементарность каждого нук-леотида матрице дважды: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний ( З - концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка ( например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью которая отщепляет не спаренный с матрицей нуклеотид затравки, после чего полимеризация восстанавливается. Этот механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Мутации, нарушающие З - экзонуклеазную активность ДНК-полимеразы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной активности относительно полимеризующей, снижают темп мутирования генетического материала. [56]
ДНК-полимеразы проверяют комплементарность каждого нук-леотида матрице дважды: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний ( З - концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка ( например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью, Которая отщепляет не спаренный с матрицей нуклеотид затравки, После чего полимеризация восстанавливается. Этот механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Му-Тации, нарушающие З - экзонуклеазную активность ДНК-полимера-Зы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной активности относительно полимеризующей, снижают темп мутирования генетического материала. [57]
ДНК-полимеразы проверяют комплементарность каждого нук-леотида матрице дважды: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний ( 3 -концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка ( например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью, Которая отщепляет не спаренный с матрицей нуклеотид затравки, после чего полимеризация восстанавливается. Этот механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Мутации, нарушающие З - экзонуклеазную активность ДНК-полимеразы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной актив - Поста относительно полимеризующей, снижают темп мутирования генетического материала. [58]
Страницы: 1 2 3 4