Фосфатная группа
Cтраница 3
Отрицательные заряды фосфатных групп должны быть экранированы ионами солей в угоду сохранению стабильности двойной спирали ДНК. Возможно, ионы могут выступать и как репрессирующие агенты молекул ДНК. [31]
Вторая функция фосфатных групп очевидна: они являются необходимыми компонентами в процессе ферментативного запасания метаболической энергии, поскольку они в конце концов передаются на ADP с образованием АТР. [32]
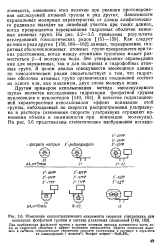 |
Изменения концентрационного инкремента скорости ультразвука при ионизации фосфатной группы в составе различных соединений [ 149, 163J. [33] |
При приближении фосфатной группы к другим атомным группам вытесняется часть воды из гидратной оболочки и эффект ионизации уменьшается. [34]
Увеличенное экранирование фосфатных групп при ионной силе выше 10 - 3М является однородным и асимптотическим, поэтому вызвать смещение р / С в сторону более низких значений можно лишь постоянно увеличивая концентрацию соли. Однако при низкой ионной силе вследствие диффузии противоионов в направлении от периферийных фосфатных групп двойной спирали появляются участки, в которых неэкранированные фосфатные группы могут - создавать очень сильное поле. Можно ожидать, что с уменьшением концентрации ионов число и размеры таких участков будут увеличиваться. Кроме того, можно ожидать, что протонирование ДНК, оказывающее влияние на прочность водородных связей и ослабляющее спиральную структуру вследствие увеличения электростатического отталкивания при понижении степени экранирования противоио-нами, будет сопровождаться разделением двух цепей и последующим разрушением структуры. [35]
Такое строение фосфатных групп, а также асимметрия карбоксильных групп у остатков серина говорят о цвиттерионной форме молекул фосфосерина в кристалле - - НОзРОСЙ2СН ( КНз) - СООН. [36]
Характерно наличие фосфатной группы между двумя сахарными остатками. [37]
Необходимость присутствия фосфатной группы для проявления активности наиболее просто интепретируется, если допустить, что эта группа действует как рычаг для осуществления индуцированного напряжения в фермент-субстратном комплексе, однако возможны и другие интерпретации этого явления в терминах гипотез индуцированного соответствия или непродуктивного связывания. [38]
Возможность миграции фосфатных групп в кислой среде у миоино-зитмонофосфатов и миоинозитполифосфатов определяется цис - или транс-ориентацией соседних гидроксилов. Так, перемещение фосфатного остатка к соседней ц с-гидроксильной группе идет в значительно более мягких условиях, нежели к трамс-гидроксилу. Миоинозитполифосфаты, содержащие одну из фосфатных группировок в положении 1 или 2, быстро изомеризуются при нагревании с кислотами, эфиры с иным положением фосфатных остатков в этих условиях значительно более устойчивы, а миграция фосфатного остатка к соседней гра с-гидрок-сильной группе затруднена. Периодатное окисление продуктов кислотной изомеризации миоинозитполифосфатов дает возможность дифференцировать миоинозиттрифосфаты, содержащие фосфатные группы в положении 1 или 2 и отличающиеся местом прикрепления остальных фосфатных остатков. [39]
Для обнаружения фосфатных групп РНК, связанных с белками, материал фиксируют смесью Кар-нуа в течение 8 - 10 часов или 5 - 10 % - ным нейтральным формалином в течение 12 - 24 часов. Фиксированные смесью Кар-нуа объекты обрабатывают дважды абсолютным этанолом ( по 12 часов), затем проводят через промежуточные среды по общепринятой гистологической технике и заливают в парафиновые блоки. [40]

Расходование двух фосфатных групп АТР на активацию одной молекулы предшественника может показаться расточительством, т.е. может сложиться впечатление, что энергия фосфатных групп расходуется в этом случае неэкономно. Позже мы увидим, однако, что за этим кроется важный механизм, обеспечивающий завершение некоторых биосинтетических реакций. [42]
Поэтому взаимодействие соседних фосфатных групп одной цепи и в нативном, и в денатурированном состоянии должно быть примерно одинаковым и не должно оказывать существенного влияния на различие в стабильности этих двух состояний. Однако взаимодействие между удаленными фосфатными группами, принадлежащими одной и той же цепи, может быть различным в нативном и денатурированном состояниях вследствие различия конформаций двухспиральной и односпираль-ной молекул. Если же этим эффектом пренебречь, то следует принять, что основным различием между двумя состояниями является отсутствующее в одноцепочечном полинуклеотиде взаимодействие между фосфатами, принадлежащими противоположным цепям двухспиральной структуры. [43]
Чтобы ввести фосфатную группу в конец, олнгонуклеотидной цепи ( для получения промежуточных фосфатов, необходимых в синтезах типа описанных выше), обращаются к обычным методам фогфорилирования, применяя их к соответствующим образом защищенным соединениям. [44]
О в фосфатной группе равно 1 61 О. [45]
Страницы: 1 2 3 4